In a series of papers published last few decades, we offered data and explanation that variation of quantitative characters could not be explained only on the basis of classical hypothesis of cumulative effects of individual loci , but, in addition, on the necessary balances of homo- and heterozygous pair- relationships in corresponding groups of polygenes. It correlates with a cumulative effect of homo-vs. hetero- combinations of corresponding allelogenes, that contribute to a more efficient reduction from a huge theoretical, to an adaptively restricted and limited variation.
Anthropogenetics; qual./quant; Characters; Darwin/Mendel; HRCtest
Table 1 presents a model of Mendelian (‘free combinations’) and Darwinian (‘natural selection’) variants, as determinants of quantitative traits, in a process of reduction to adaptive variation, based on a cumulative effect of individual loci [1-3], versus such effects of the ratios of allelic homo- and heterozygosities [4-5]. This second turns out to be more efficient, with the only possibility to explain the maintenance and inheritance of variation among the progenies of survived parents from peripheral classes. Whichever ‘Darwinian class’ survives in the nature, it provides almost equal variability of ‘Mendelian variants’ in this and following generation, with a possibility of adaptive reduction which is many-fold in comparison with those with classical ‘additive-genetic’ hypothesis. The basic unit of Darwinian selection are not individual alleles, but pairs of alleles, that determine which combinations at chromosomes will mach and survive in meiotic cell divisions. Quantitative-genetic variabilities exist among ‘Darwinian’ classes with specific degrees of homo/heterozygosity, as well as within the colons of these classes (‘Mendelian’ diversity) that provide an ubiquitous capacity for variation of their progeny, whichever class survives from parental generation.
Table 1. Quantitative traits genetically controlled according to cumulative effect A/ of additive genes, or R/ of their homo- versus heterozygous ratios. The progenies of F1 hybrids (AaBb x AaBb, etc.). Diversity among horizontal classes belongs to phenotypic ‘Darwinian variability’, and within vertical collons it is ‘2nd law Mendelian variation’.


Using as a model the classical investigations of homozygous genetic loads in Drosophila from 1960s–1970s, we decided to observe homo-heterozygous relationships in an analysis of recessive quant. and qual. variants of human anthropo-genetic properties. 20-30 observed and strictly selected homozygous- recessive characters of humans (Table 2) were compared in different samples of studied individuals, with the aim to visualize the nature of homo- and heterozygous relationships, and amounts of genetic loads that are eventually present in carriers of studied samples. Out of tens of such comparisons in last three decades, e.g., between samples from hospitals and controls [6-10], we present here just a few.
Table 2. Test HRO / HRC-TEST*
Broj osobine |
Homozigotno-recesivne
osobine Homo-recessive characters |
Broj osobine |
Homozigotno-recesivne osobine
Homorecessive characters |
1. |
Plava kosa - svetla boja kose
Light hair |
12. |
Nesposobnost uzdužnog savijanja jezika
Inability to curve tongue vertically |
2. |
Prava kosa
Straight hair |
13. |
Nesposobnost rolanja jezika
Inability to curve tongue longitudinally |
3. |
Dva cveta u kosi
Two roots in the hair |
14. |
Desni palac preko levog
Right thumb in clasping over left thumb |
4. |
Obrnut cvet u kosi
Opposite root in the hair |
15. |
Savijanje palca pod uglom > 450
Distal part of thumb curves back more than 45 d. |
5. |
Mekana kosa
Soft hair |
16. |
Hiperekstenzibilnost palca Tumb hyperextensibility |
6. |
Prava linija kose na čelu
Straight line in front of the hair |
17. |
Ubacivanje zglobnog palca
Thumb proximal incorp. movability |
7. |
Vezan ušni režanj
Joined ear lobe |
18. |
Tri žile u korenu šake
Three tendons in ventral join of the arm |
8. |
Uho bez Darvinove kvržice
Ear without Darwinian knot |
19. |
Levorukost
Lefthandedness |
9. |
Plave oči
Blue eyes |
20. |
Indeks prstiju (duži ili isti II od IV prsta kod muškaraca
i duži ili isti IV od II prsta kod žena). 2nd finger equal or
longer than 4th in males, opposite in females. |
10. |
Daltonizam
Daltonism |
21. |
Ne oseća ukus PTC-a
Nontasters of phenilthiocarbamide. |
11. |
Grleno »r« ; Gutural 'r' |
|
|
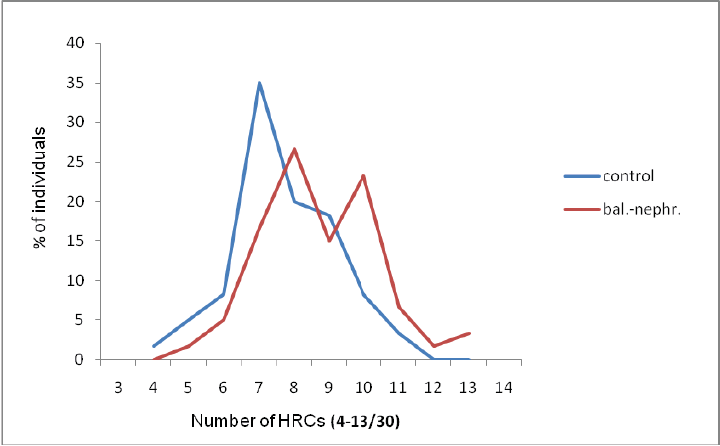
Balkan Endemic Nephropathy
HRC-test in healthy individuals and patients from place Niš and Aleksinac
Chronic Obstructive Pulmonary Disease
Among 150 patients of COPD no. HRCs is signif. increased
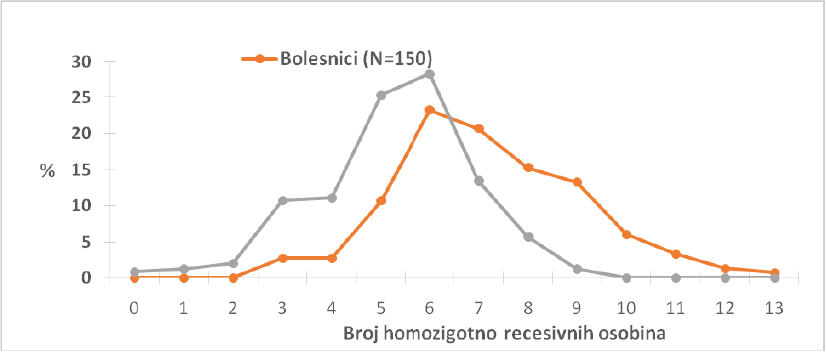
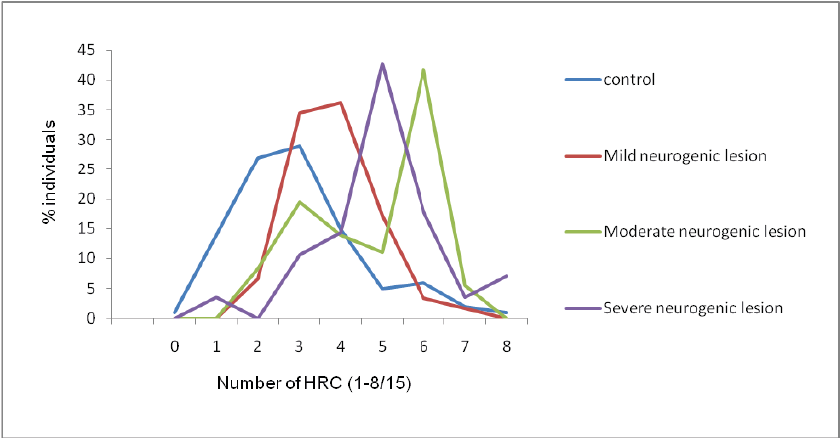
Spina Bifida
Frequencies of homozygous recessive characteristics (HRC) in different groups of patients with spina bifida and control.
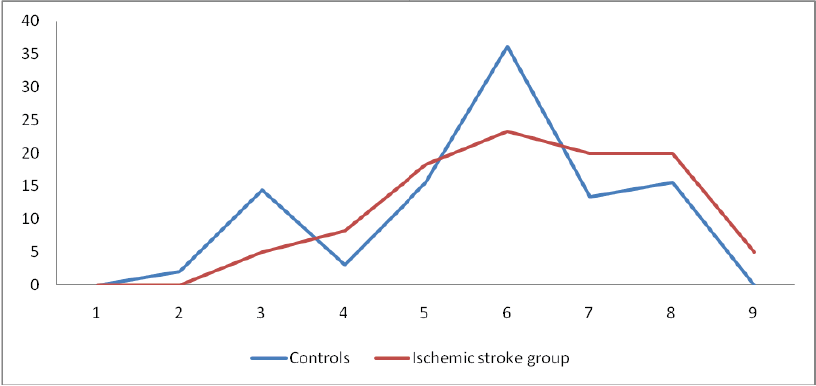
Ischemic Stroke
Frequencies of homozygous recessive characteristics (HRC) in controls and in manifested ischemic stroke patients are different
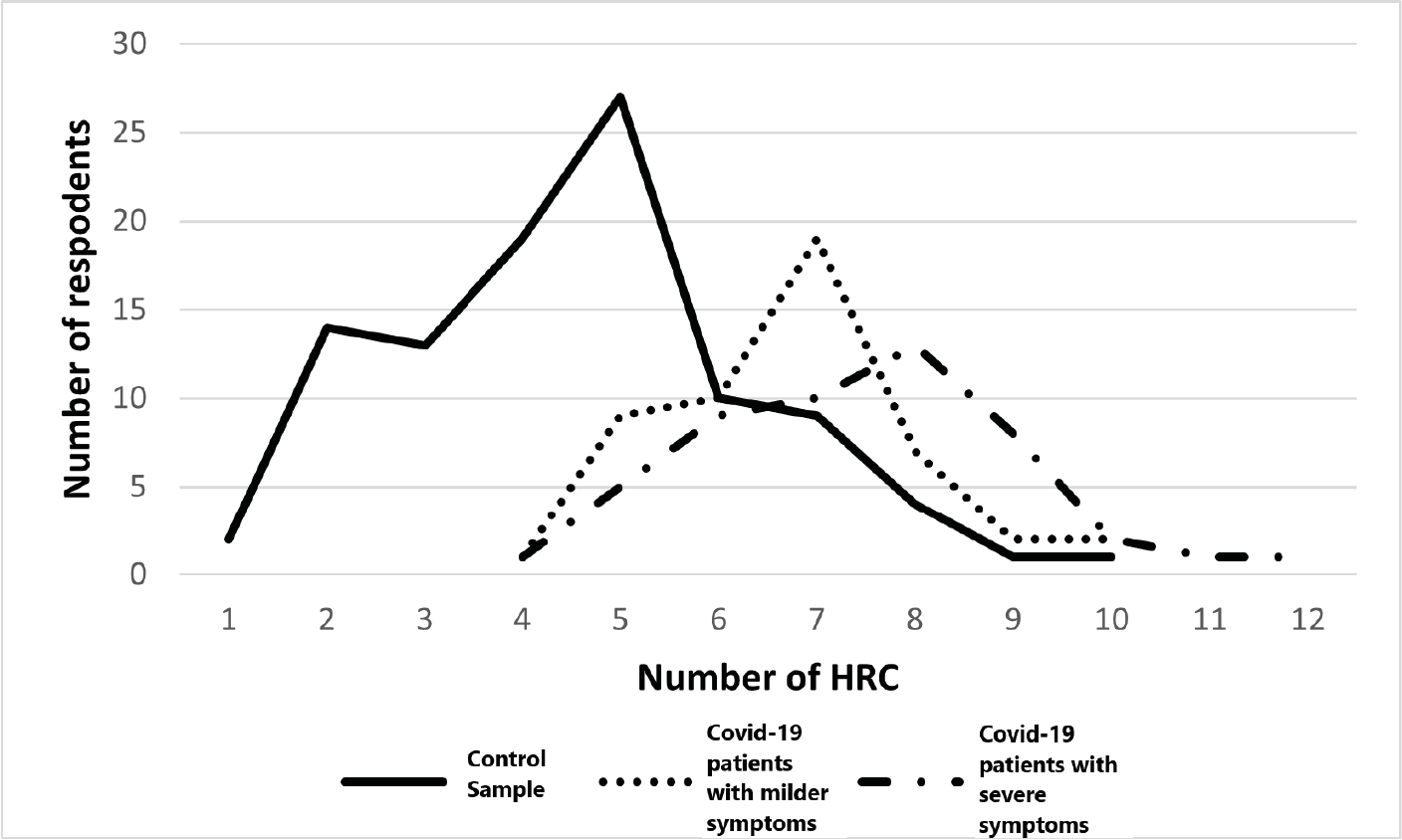
Covid-19
HRC-test
Determination of quantitative traits by homo / heterozygous balances gives the same ratios in progenies of F1 hybrid parents as in long approved theory based on ‘additive genes’ control (Tab.1). The ratios of phenotypes for characters determined by 2, 4, 6 and 8 pairs of alleles correspond to the kinds of ratios obtained in numerous experiments explained by ‘additive genes’ at 1, 2, 3, and 4 loci, respectively. Differently from ‘additive genes effects’, homo/hetero genetic balances provide approximately equal allelogenic constitution in all classes that may individually survive under specific ecological conditions, enabling a quick recovery of genetic variability in following generations of a population. The difference is especially crucial when one of peripheral classes survives, and ‘additive genes theory’ does not give a solution how this reconstitution of biol. variability may occur. Huge Mendelian variability, based on 2nd rule of free combinations of parental gametes, cannot be adaptively reduced if we exclusively use ‘additive genes theory’ for determination of quantitative traits. Instead of 16 in A’’, even 256 Mendelian variants (in R’’ from Tab.1) could be reduced to five Darwinian ‘channels’ if we apply ‘balanced theory’ of determination of quantitative traits [4, 11, 12]. With a ‘cascade’ of such processes before and during gametogenesis, we approach a solution of so far enigmatic question what are the origins of a fascinating similarity of individuals within everyone of organismic species [12,13].
During passed three decades in Chairs of genetics at the Faculties of Biology and Medicine in Belgrade, the analyses are proceeded of a series of qualitative anthropogenetic characteristics to determine the degree of genetic homozygosity (HRC-test) in samples of humans with different but specific adaptive properties. It has been determined that such properties could be indicators of genetic loads that exist in specific groups of individuals, carying also determined predispositions of specific adaptations. An increased degree of homozygosity is in a majority of cases present among patiens from clinics visited for such analyses, such as in cardiovascular, urogenital, neuropsychiatric, part of ortopedic clinic, but also among alcoholics, and in a milder degree among lefthanded persons and female smokers. Extremely low HRC-values are determined among sportsmen, as well as in samples of homosexual individuals [9]. It is of special value to analyse the similarities between different populations, from so called 'normal' samples with the average HRCs. Application of HRC methodology is in a possibility to predict some developmental properties of young individuals, with a positive control and better understanding of their maturation and development. All described dispositions must not have a common root and growth origin, but they do contribute to a physiological balance that results in so called 'adaptive combination' of fitness properties in humans.
- Nilson-Ehle H (1909) Lunds University Aarskr. N. F. Afd 2: 5-122.
- Emerson S., East EM (1913) The inheritance of quantitative characters in maize. Nebrasca Agric. Exp. Res. Bull.
- East E.M (1916) Studies on size and inheritance in Nico- tiana. Genetics 1, 164 1:164.
- Marinkovic D. A (1977) new model explaining the variation of quantitatative traits. GENETIKA 9: 17-27.
- Petanović R., Marinković D (1978) Experimental proof of classical theory and a new model of quant. traits variation. GENETIKA 10: 151-164.
- Marinković D., Spremo B., Ilic M (1990) Studies of human population-genetic variation in special and regular schools in Serbia. Arch. Biol. Sci 42: 11-12.
- Marinković D., Jovanović D., Cvjetićanin S., Savić M., Branković S (1994) Genetic homozygosity and normal vs. pathological human variation. GENETIKA 26: 147-156.
- Marinkovic D., Cvjeticanin S (2013) Anthropogenetic Ho- mozygosity and Adaptive Variability. Serb. Acad. Sci. and Arts, Monographs, book 172: 1-215.
- Marinkovic D., Cvjeticanin S (2020) Genetic balances that maintain potential hypo and hyper-activities in physiologi- cal and behavioral characters. Int. J. Res. Studies in Med. Health. Sci 5: 1-3.
- Petronic I (2020) Morphogenetic variability as potential biomarker of neurogenic lesion degree in children with Spina biphida. Healthcare 8, 68 2020. Book chapter in: Chalenges in Desease, Health Research 1-9.
- Marinkovic D (2016) Two crucial approaches in contempo- rary developmental and evolutionary genetics. Int. J. De- vel. Research 8351-8354.
- Marinkovic D (2017) Genetic balances as determinants of biological similarity and variation. Int. J. Devel. Research 7: 15283-15285.
- Grujicic D., Milosevic-Djordjevic O (2021) HRC biomarkers of predisposition for Covid 19. Genes and Diseases, in press.